Cortical subcortical integration and decisions: A prefrontal cortex – nucleus accumbens neural circuit case study
Vanessa Nyblom UA ’25 and Jim Stellar
In our last blog, we consider the prefrontal cortex (PFC) interactions with the amygdala as part of the negative (fear) side of a subcortical system that could contribute to cortico-limbic decision making. While that last blog focused on things that the organism should avoid (e.g. pain, fear), here we focus on the positive side (reward, pleasure) and things the organism should approach. But the story of cortical-limbic interactions is similar, only this time the PFC connects to the Nucleus Accumbens and the Ventral Tegmental area (VTA). These two systems (PFC-Amygdala and PFC-accumbens/VTA) could be the “guardrails” for emotional/limbic input to cognitive decision-making.
Our story with a the PFC-accumbens/VTA interaction starts with glutamate projection from PFC to both the VTA and the accumbens as diagrammed below. Remember, according to classic reward neuroscience, it is the dopamine neurotransmitter release from the VTA and particularly into the accumbens that underlies the reward experience in rats and humans.
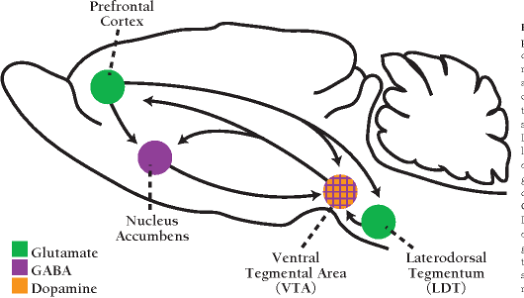
In this circuit, taken from the rat, the PFC uses glutamate to interact not only with the accumbens but with the source of dopamine that comes to the accumbens from the ventral tegmental area (VTA) and is thought to represent the reward experience itself. Like the last blog, there is a reciprocal neural circuit connection from the VTA back to the PFC. Unlike the diagram in the last blog, the PFC projections also go to the Laterodorsal Tegmentum which has a well-known connection to the VTA. The general idea is that through these connections the PFC can regulate the positive emotional experiences coming from the accumbens through the VTA, just like it can for the negative emotional experiences in the PFC-amygdala ciricuit.
How might this work? To begin, we go back to another blog featuring an important older paper where the VTA neurons in the monkey were studied in terms of their firing pattern in reaction to a reward delivery. What was concluded in that blog was that the VTA neurons were “smart,” as shown in the following diagrams from the original study and discussed below the diagram. In this diagram the authors plot the VTA firing temporally as a reward is given and as conditioned stimulus (CS) training is applied.
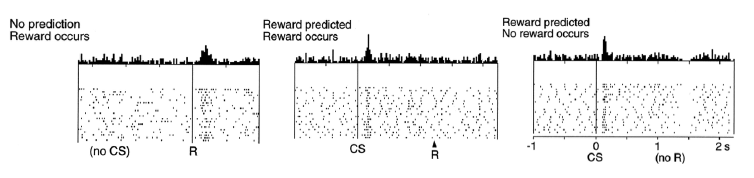
Here is how the diagram works: In the left diagram, the VTA neuron is seen to fire every time when a reward is given. In the middle diagram, when the CS is established through Pavlovian learning, the VTA neuron fires to the CS,. But somewhat surprisingly it remains at its baseline firing rate when the reward is delivered. One is left to think from this second result that what matters to the VTA neuron is that the reward is certainly coming, and if that is known, the actual reward itself may not ne as important. Finally, in the right diagram, when the reward is omitted the VTA “notices” by going below baseline activity exactly when the reward should have appeared, even though it still fired to the CS as usual. This particular response was seen as the reward prediction error that signals that the organism needs to change its behavior because something is not working. Note there are more recent studies that are even more sophisticated.
Now, with the addition of this PFC glutamate pathway to this thinking, perhaps we can revise our ideas to conclude that it is not the VTA that is showing this “intelligence” but instead the PFC is “playing the VTA neurons” like a pianist plays the keys of the piano. That may indeed be the case for the accumbens core neurons that receive VTA input.
In any case, the PFC-accumbens/VTA regions are entangled together in reciprocal neural circuitry, just like they are in the amygdala blog. And that is the main point of these two blogs – that what underlies decision-making contains both cognitive and emotional components, and these two systems serve as the limbic guardrails for cognitive decision. However, really it is the opening to further investigation of cortical-limbic integration.
But before we stop we need to mention one more interesting point to make about this glutamate pathway, and that is the VTA also releases glutamate.
The Glutamate Pathway from the VTA (not just dopamine)
In an interesting recent paper (that reviews another paper published in the journal Neuron), glutamate is discussed as being released from the VTA into the accumbens, not just from the PFC. That adds another level of complexity to this story. In the diagram below some of VTA neurons are shown in blue that co-release glutamate and dopamine into the accumbens.
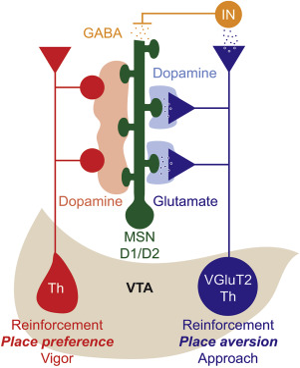
What makes this line of work interesting is that using other molecular biology methods the original paper authors were able to delete the dopamine by genetically blocking the function of tyrosine hydroxylase (TH in the diagram) but not glutamate. They then showed in a classical electrical self-stimulation reward behavioral paradigm that mice still found the stimulation rewarding. In an even more interesting finding they showed that stimulating this pathway produced in a classical place-preference reward behavioral paradigm produced the opposite. Mice showed place-avoidance or aversion.
What does this result tell us? How can the VTA be in charge of aversion which shapes behavior away from stimuli not toward them as reward does? This adds to the story of why the PFC and VTA have a glutamate pathway between them – to allow for this feeling to be the product of both and not just the VTA. Notice in the above diagram the VTA seems to suggest that the motivation that we have for a reward-associated stimulus, may be different from the pleasure we have once a task is complete and a reward is successfully obtained.
We can also look closer at the neurotransmitter in the VGluT2 system, shown in the figure above and discussed in the paper, and notice that it meditates dopamine storage within the vesicles of the accumbens. In doing this, we can see that it controls the glutamate-dopamine releases. We can infer that this release, and the VGluT2’s role in it, affects the motivation effect on one’s brain. As seen in the figure, the reinforcement, whether it be through place preference or place aversion, is brought to the VTA through the dopamine, glutamate, and GABA neurons. We know that the VGluT2 system in the nucleus accumbens in that diagram just above uses glutamate entry to enhance storage and co-release, showing that this is coming from the dopaminergic parts of the accumbens, and could mean that the reinforcement of motivation is coming from those same neurons firing in the accumbens.
Conclusion
To paraphrase the old political slogan of “It’s the economy, stupid.”, we would say here that “It’s the loops stupid.” While this does not help us understand the deep and specific connections between the amygdala and the PFC or the accumbens and the PFC, and it does not help us understand how the PFC brings emotionally negative and positive input into doing its job of planning and emotional regulation, it is a start. Recurrent neural circuit loops also are a favorite device of the nervous system for behavioral regulation and computation. Finally, we need to look at how the PFC as a special 6-layered cortical region (and other brain areas such as the anterior cingulate cortex or the insular cortex) represent these emotions inputs from the limbic system so they can enter the value proposition. Remember, this blog is about how learning from experience in college complements the learning of facts and theories from the classroom to make students more mature and even wiser about their chosen fields of study. These mechanisms are probably at work here even if we tend to be more aware of the explicit learning from the classroom than the implicit learning from the direct workplace or other experiential site of knowledge application.