Symbolic Representation in the Neocortex: White matter and the dynamic cortical networks
By Kassie von Stein UA’25, Vanessa NyBlom UA’25, Onoseta Momodu UA’24, Pauline Meunier PS’23, Rachel Orenstein UA’24, Brandy Eggan, and Jim Stellar
In a previous blog, we wrote about how repeating neocortical columns are an unusual feature of the nervous system’s evolutionary development. They are extensive in mammals and especially in primates. We argued in that blog that this columnar structure developed to support a cognitive symbolic logic process not seen in limbic system, motor system, and other important lower neural processes. That cortical columnar system increases the organism’s ability to interpret and deal with the world. In another blog, we discussed how such a neocortical system could re-represent the fear-type emotional processing of the brain’s own limbic system to add a value component, such as the idea of risk, to the brain’s cognitive decision-making and planning. Work there is ongoing. All of this cognitive-emotional processing goes back to the general theme of this blog series, which explores the brain basis of how college students mature from direct experiences like internships within their academic fields. Finally, we note a recent finding from the connectomics field that shows the incredible complexity of the human cortex, with which we will not deal in this writing.
In this blog, we specifically look at the neocortical connections between columns as they go between neocortical regions. These connections are often called “white matter” because of the appearance of the myelin surrounding their axons. It is different from what is called “gray matter” in the cortical columns where computations occur. These white matter connections between columns are an important part of the symbolic processing by the neocortex. They work to connect what may be separate cortical processing functions in lower animals. Also, white and gray matter clearly operate together in relevant networks that can be dynamically switched on or off as tasks demand. So to end, we will look briefly at neural networks, e.g. the Default Mode Network, that are made out of these pathways and how they may be activated or not as circumstances change.
The development of white matter in evolution
Parallel to our previous blog, we note that white matter connections in the cortex also expand with evolution (as seen in the graph below).
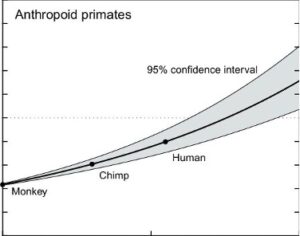
This development may not be surprising, as one would expect white matter to expand along with the neocortex itself. But this expansion also happens in conjunction with the greater ability of humans, even over primates to do integrative tasks. An impressive such task is cross-modal transfer between object representations.
The example of white-matter-based sensory cross-modal transfer of object representations
Consider the moment when you are searching for your keys, and then feel them in your pocket. You then have an immediate visual image of how the keys appear – almost as if you could see them through the fabric of your pocket. We humans do this effortlessly, but it is not so in other animals – with the exception of some primates in a study of visual-tactile transfer where they were trained to use symbols to communicate. In an earlier study, untrained monkeys could not consistently make that transfer, and the evidence of cross-modal transfer in an audio-visual domain is even weaker in cats and dogs.
In humans, the topic of cross-modal transfer has received consistent attention from neuroscientists. In a 2003 study of the underlying brain circuitry of cross-modal transfer, which combined brain activity with fMRI (functional magnetic resonance imaging) and brain anatomy MRI (magnetic resonance imaging), researchers studied cross-modal transfer pathways between touch and vision. These authors even identified a brain area with fMRI, the posterior intraparietal sulcus, that was activated in their experiment’s cross-modal transfer task. A type of brain scan called “diffusion tensor imaging” can measure these axonal connections directly, and such a study in 2011 revealed direct white-matter connections between tactical and visual areas. In a more recent 2017 study, another group of subjects was also observed to have this ability, though with some variability that was linked to differences in the microstructure of the white matter connections themselves.
The visual system is known to divide its processing into a ventral visual pathway that is associated with information, such as objects, and a dorsal pathway that is associated with movement and action. According to a 2015 paper, tactile inputs even seem able to activate visual areas not only in the ventral pathway for objects, but also in the dorsal pathway based on movement and action.
An interesting but usual case in a 2020 paper, people had their sight restored after living in a world where their primary distance-sense was hearing. This study was done by taking advantage of a new gene therapy that restored sight in a small group of patients who were studied for years before and after restoration. Before, these patients had robust responses in the occipital cortex to auditory input. That response continued after sight restoration. It is interesting to think about what the symbolic representation of auditory and sensory stimuli are in these patients, and how their experience differs from an average person. We know that sensations can cross, causing the phenomenon of synesthesia, but that topic is also beyond the scope of this blog, albeit intriguing.
The implications for the representation of white matter connections across sensory areas are huge. For example, your representation of a familiar complex stimulus like your cat may be very multidimensional to you, consisting of sight, sound, feel, etc. But the representation your cat of you is likely to be somewhat less so. With considerably less of the white matter than you and other primates have to interconnect sensory neocortical brain areas, we can guess that the cat only fully perceives you as multidimensional entity when you are present. You bring all of these different sensory representations to the moment. What the cat carries in its head when you are not around, and this is purely speculative, might be separate streams of your stimulus presentation. That may be enough as cat-human interactions are clearly robust.
The question for humans is whether this multidimensional processing extends to symbolic representation of limbic processes – about which we write in this blog. We have said that emotion brings value to cognitive knowledge and maybe it plays a role in the cat story above. But how? One way to consider that here is by looking at networks. Then we can return to the fundamental problem of cognitive-emotional integration and white-matter connections.
Neural networks starting with the Default Mode Network
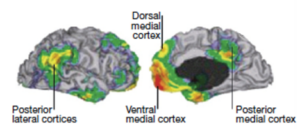
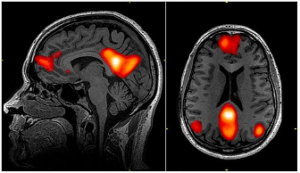
The first thing one needs to know about neural networks is that they can interconnect wide-ranging cortical areas and they can change rapidly. Consider the well-known Default Mode Network (DMN). As we have written previously, the DMN seems to connect the front part of the brain, where motor function and importantly planning is organized, with the back part of the brain, where sensation is processed and objects and their actions are represented. It is shown in the left-side diagram below, taken from an fMRI brain activity paper where the outside of the brain is shown on the left and the inside of the brain is shown on the right. In this study the subject was asked to do nothing, but, as is typically the case, engaged in mind-wandering. It was this default (nothing to do) condition that gave rise to the name of this network. The DMN is so popular that it is even featured in Wikipedia, which we cite here if only to make the point of its widely known status. We also use that same link to show a diagram on the right-side below. It shows the same DMN brain area activations, but it is taken directly from fMRI images and features a view from the side of the head (left) and a view from the top of the head (right).
The idea here is that the white matter connections shown in the diagram, below support that link. This figure came from a diffuser tensor imaging (DTI) brain scan and was also from Wikipedia. It shows the white matter connections between the front and back of the brain the front being on the left side.
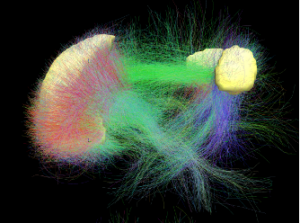
More centrally, these connections, while always present, are not activated until the circumstances call for it. For example, if you are doing something, a Task Positive Network (TPN) is activated and the default mode network is not. In fact the TPN and the DMN may be antagonistic networks where only one can be operative at a time.
In an interesting aside, this interaction was first seen by JS when he heard a talk while in graduate school at the University of Pennsylvania in the late 1970s by David Ingvar. In that talk, he remarked on the observation that when a subject was asked to wait quietly in a PET (positron emission tomography) brain scan, the frontal cortex had enhanced blood flow. When Professor Ingvar asked the patient what he was doing, JS remembers him saying that he was thinking. It was not until the early 2000s, some 30 years later, that the term DMN was coined and it burst onto the neuroscience scene.
The TPN and the DNM are two of what we now think are 6 or more commonly accepted brain networks. They go by different names and some imply different functions. For example, in the list above the TPN seems to be the “salience network” which is not quite the same thing. A modern understanding of brain networks is beyond the scope of this blog. It also potentially leads to a discussion of consciousness, which is again beyond the scope of this blog.
Conclusion
The point of this blog follows the general direction of this blog series in long and short forms. That direction is to explore the potential brain mechanisms of how in college students some direct learning from experience such as an internship that often occurs outside the classroom is a brain-natural way to make the academic curriculum more powerful in terms of student career-type decisions. By doing that we hope to better understand the integration of what we think of as older brain processing in the limbic system with the more recently evolutionarily developed neocortical system with its two outstanding characteristics: The repeated use of an impressive number of cortical columns, discussed in a prior blog, with the massive interconnections between those cortical columns from many specific processing areas as introduced here as sensory areas that somehow connect to produce cross-modal transfer and true multi-modal or poly-sensory representations in humans. How that combines with limbic input, perhaps to bring value calculations into cognitive decision-making, is the subject of other blogs and goes back to the cognitive-emotional or cortical-limbic integration that we think is necessary to promote growth in college students so that they develop to the best extent possible at least the beginning of what we like to call professional wisdom.