Symbolic Representation in the Neocortex: Which brain areas represent which limbic processes and how do they work together?
By Onoseta Momodu UA’24, Vanessa NyBlom UA’25, Kassie von Stein UA25, Pauline Meunier, PS’23 Rachel Orenstein UA’24, Brandy Eggan, and Jim Stellar
Introduction:
This blog begins to examine neuroscientific literature on the complex relationship between the limbic system’s emotional (somatic) markers and the neocortical system’s cognitive decision-making processes, primarily focusing on the frontal cortex.
In this exploration of the cognitive neurobiological basis of decision-making, we highlight the input of two key limbic areas, the amygdala and nucleus accumbens, representing negative and positive emotional reactions. We suggest that their input is handled by the cortex at a symbolic level for processing and planning and we focus on five key cortical areas: the orbital frontal cortex, insula cortex, ventromedial prefrontal cortex, anterior cingulate cortex, and dorsal medial cortex. By discussing these areas and their interactions, we hope to begin a more nuanced understanding of how positive and negative reactions and emotional somatic markers intricately shape our choices.
The blog continues the exploration of the neuroscience behind multifaceted mechanisms governing the impact of emotion on cognitive planning and decision-making. This work fits with the ongoing blog series application of how direct experiences impact college students planning potential career paths. In previous blogs, we wrote about general symbolic representation in the neocortex due to a unique cortical column structure and the columnar white matter interconnections in networks. Finally, this blog post opens up a return to the individual brain areas discussed here and in previous shorter blogs, but in greater depth, really as more through long blogs. One of those key cortical areas discussed is in another long blog on the insula cortex and what we think is the re-representation of fear and fear conditioning as a more abstract and cognitive process of risk and risk management.
At this point, we will step through the five cortical areas mentioned above and incorporate two side notes on approach and withdrawal behaviors as part of limbic structure as well as a side note on the somatic marker hypothesis that seems particularly relevant to the insula cortex.
Orbital Frontal Cortex
weighing alternatives
As we previously wrote about in a blog, the orbital frontal cortex (OFC) is situated in the frontal lobes above the eyes. The diagrams below, taken from that blog, depict connections with limbic system circuitry, particularly the nucleus accumbens on the left and the amygdala on the right.
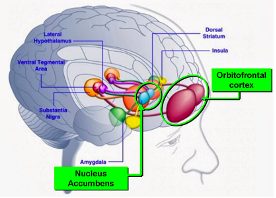
The OFC holds a central role in decision-making and executive functions. Its involvement in decision-making is multifaceted, encompassing the evaluation of potential outcomes and consequences, as well as the selection of optimal choices based on the assessment of rewards and punishments. It also coordinates executive functions such as planning, working memory, and inhibitory control, facilitating adaptive and goal-directed behavior. Here the OFC, working with the frontal cortex in general, acts as an integration hub for emotional and cognitive processes.
The OFC’s inputs and outputs reflect emotional input information to cognitive decision making from both the fear and withdrawal processing of the amygdala and the reward and approach processing of the nucleus accumbens. According to some thinking, the lateral OFC in particular seems to have connections to those areas associated with punishment or failed delivery of rewards whereas the medial OFC is more associated with the processing of reward and positive outcomes. The OFC has connections with other areas, such as the dorsolateral prefrontal cortex (DLPFC) and the ventromedial prefrontal cortex (vmPFC), both of which are also involved in planning and decision-making processes. These and other connections are indicated in the diagrams above and the vmPFC in a section below.
The significance of the OFC appears to lie in evaluating the emotional meaning of choices, especially in processing information related to potential rewards and punishments. This evaluation extends to social decision-making, where the OFC aids in interpreting emotional cues and social dynamics. Overall, the OFC’s nuanced contributions underscore its importance in forming somatic markers—bodily responses associated with emotions that act as quick, unconscious signals influencing decision-making. Somatic markers serve as physiological responses linked to emotions, such as fear, anxiety or excitement, playing a pivotal role in decision-making. These markers, often manifested as visceral sensations or gut feelings, offer quick and intuitive assessments of potential outcomes associated with choices. The significance of emotional somatic markers lies in their ability to guide decision-making, aligning choices with emotional well-being and past experiences. We are working on a long blog on the OFC building on this blog and the earlier one first mentioned in this section.
1st Side note: The negative and positive underpinnings of the amygdala and the accumbens
The amygdala is part of the limbic system located deep in the temporal lobe. It contributes to the rapid appraisal of emotionally negative stimuli, forming associations crucial for the representation of somatic markers. The nucleus accumbens, located in the anterior portion of the ventral striatum, plays a role in assessing the rewarding aspects of choices through the release of dopamine. Of course there are many interactions between these systems, such as a ventral tegmental area’s dopamine projection that innervates both amygdala and accumbens as well as the frontal cortex. As an example of the complexity of this circuit, the frontal cortex sends projections back to both the amygdala and the accumbens.
The amygdala is recognized for its swift assessment of negative environmental cues. It rapidly distinguishes emotionally significant stimuli, particularly those bearing potential threats or rewards. Notably associated with the fear response, this structure contributes to the rapid activation of physiological reactions in response to perceived dangers. Furthermore, the amygdala actively participates in the consolidation of memories related to fear-related emotional experiences, reinforcing the enduring impact of such events. It also links emotional valence, denoting the positive or negative nature of emotional experiences, to decision-related neural networks, and it influences the cognitive appraisal of situations, shaping subsequent actions and affecting risk perception. Through its role in emotional conditioning and learning, the amygdala forms associations that prove crucial for the generation of somatic markers – physiological responses associated with emotions. Dysfunction in these processes may contribute to impaired decision-making, as observed in conditions like post-traumatic stress disorder (PTSD) or anxiety disorders. The amygdala’s intricate interplay in emotional processing, associative learning, and decision-related neural networks underscores its profound impact on the emotional and cognitive dimensions of decision-making particularly in fear/anxiety but also more broadly.
The nucleus accumbens, a pivotal component of the brain’s reward system, plays a fundamental role in processing rewarding stimuli and mediating the experiences of pleasure and reinforcement. Neurons in the nucleus accumbens are impacted primarily through dopaminergic pathways originating from the VTA. This pathway serves as crucial mechanisms underlying the ability of dopamine neurons to support instrumental learning. That is the release of dopamine in the nucleus accumbens, resulting from positive outcomes related to decision-making, reinforces behaviors associated with those outcomes. This process contributes to the formation of habits and the motivation to repeat actions perceived as rewarding. The reciprocal connection between the nucleus accumbens and the orbitofrontal cortex (OFC), already mentioned, further enhances the integration of emotional and cognitive aspects of decision-making. Dysregulation in these neural circuits can lead to difficulties in evaluating the emotional significance of choices, potentially contributing to impulsivity or impaired judgment. Understanding the dynamic interplay between these brain regions sheds light on the neurobiology of decision-making, providing insights with implications for various fields, including neuroscience, psychology, and psychiatry.
Insular Cortex
turning fear into risk
As we previously discussed in a long blog, the Insula cortex is buried in a deep fold in the cortex between the frontal and temporal/parietal lobes, as shown in the diagram below on the left. Also shown below on the right is a diagram from a paper on the somatic marker hypothesis discussing the input from the body sensations to the Insula cortex.
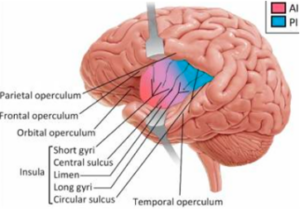
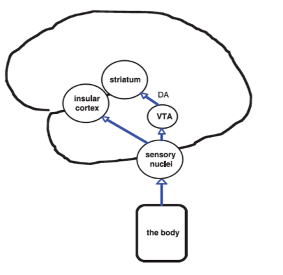
For the purpose of this blog, we are considering it along with clearly frontal cortex areas like the OFC. It is certainly part of the cortical system of reading or re-representing limbic functions, particularly the amygdala where it seems to be involved in fear and turning that into risk at a cognitive level. As previously mentioned in an analogy elevating the simple eye tracking of objects in the superior colliculus to the concept of an object having a trajectory in the neocortex, we propose that the Insula cortex is involved in elevating risk for cognitive decision-making from a Pavlovian-conditioned fear response out of the amygdala.
As depicted in the diagram above, the Insula is divided into regions, primarily Anterior Insula (AI in the diagram) and Posterior (PI). The AI-PI continuum also features a change in the number of granular cells (small neurons), particularly in cortical layer 4 with the posterior being more granular. This AI-PI distinction may be important as, for example, the AI appears in a rat study to be more tied to the Default Mode Network. This then seems to be involved with self-referential processes and is a network about which we have also written.
The Insula cortex has interoceptive inputs coming from the body mainly into the posterior Insula. There, these lower-level inputs are processed and sent to the anterior insula, where some authors think they are integrated with higher cortical regions using input from subcortical regions like the amygdala and cortical regions like the anterior cingulate gyrus and the dorsolateral prefrontal cortex. As mentioned in the original blog, the anterior insula is considered the more limbic part of the insula cortex due to these interconnections. The Insula cortex in general has output connections with the caudate and the claustrum. In the rat, it connects with a variety of brainstem and limbic structures such as the central nucleus of the amygdala, lateral hypothalamus, bed nucleus of the stria terminalis, periaqueductal gray, and parabrachial nucleus. To link to the prior discussion of the OFC, the insula projects to both parts of the OFC that we mentioned above.
Ventromedial Prefrontal Cortex
planning and the gut feeling
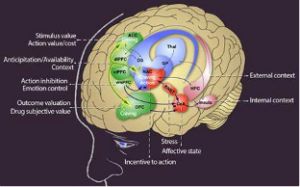
Patients with ventromedial prefrontal cortex (vmPFC) damage lack the gut-feeling to make decisions. The vmPFC is shown below.
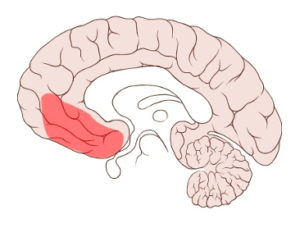
It was for this type of vmPFC patient that Damasio’s group invented in 1997 the Columbia card sorting task where subjects had to choose between four decks of cards on a screen that gave money and took it away. Before the normal subject could cognitively say that a particular deck was dangerous, the hand had a stress-related micro-sweating reaction to a galvanic skin response that could be picked up with electrodes. The vmPFC-damaged patients did not have that reaction and failed to select the correct deck. In fact, they had great difficulty making any decision, like when to schedule another doctor’s appointment. They lack that feeling of the decision. This work led to the development of the somatic marker hypothesis and further study of the card-sorting task where “gut” reactions informed conscious decisions. Damasio took this cognitive-emotional interaction to another level with his famous 2005 book, Descartes Error, where he argued that it was not, “I think therefore I am,” but more like, “I feel therefore I can think, therefore I am.” For us, consciousness is beyond the scope of this blog but the somatic marker hypothesis is not. We also recognize advances in vmPFC and somatic marker hypothesis discussion since the late 1990s and are working on a long blog on the vmPFC.
The vmPFC has inputs such as an interesting one from the posterior superior temporal cortex and inferior frontal gyrus that seems involved with social value cognition, choice, the medial striatum, the hypothalamus, and other limbic areas as well as broader connections along other frontal cortical areas. In the standard college effort of searching for a major or a career field that feels right to the student, it is no accident that the words “feels right” are used. Often this feeling is based on an experience like an internship in that career area, perhaps also coupled with some cognitive reflection after the experience to again test these feelings.
2nd Side Note: The somatic marker hypothesis
The Insula plays roles in the Somatic Marker Hypothesis, as mentioned above as being first introduced by Damasio’s group in 1990s. To go a bit deeper, this hypothesis posits that emotional information is crucial to guiding decisions as revealed by physiological indicators, e.g. the galvanic skin response in a card sorting task when the hand is over a deck with negative consequences. The idea is that when we initially make a decision about something or experience an emotionally salient event, information about the outcome is stored perhaps within the Insula in the form of somatic markers. When we face a similar situation in the future, these emotional somatic markers are recalled and integrated with cognitive information to influence what action will be taken. For example, if we are faced with a situation in which the outcome is positive, then positive feelings will be evoked while facing similar circumstances, leading to a similar decision being made. On the other hand, if the outcome was negative or painful, negative feelings will be evoked, leading to a decreased likelihood of selecting that same option.
To store an external stimulus as a somatic marker, the amygdala (AMY) acts as a trigger structure for somatic states induced by associated with the primary stimuli. They are then processed either subliminally by the thalamus or explicitly via early sensory and higher-order association cortices. Once somatic states are induced by these primary stimuli, signals are relayed to the brain, leading to the development of somatic state patterns within a circuit probably involving brainstem nuclei, somatosensory cortices (SI/SII), insula, periaqueductal gray (PAG), and cingulate cortices.
Subsequent exposure to a stimulus that evoked thoughts or memories about a specific primary inducer then acts as a secondary inducer. It is thought to reactivate the pattern of a somatic state belonging to a primary inducer, generating a fainter activation of the somatic state than if it had been triggered by the original primary inducer. If the somatic state associated with the secondary inducer is successfully encoded, future generation of somatic states from secondary inducers are dependent upon cortical circuitry, especially the vmPFC, which then acts as a trigger structure for the generation of somatic states from secondary inducers. In Damasio’s formulation neurons here and in other places serve as a convergence-divergence zone that connects wide-ranging brain areas particularly in the limbic-cortical interactions we are discussing here. These neurons are thought to couple certain categories of events based on memory from cortical areas with effector brain structures that produce a somatic state in the body (such as seen in the galvanic skin response). In essence, the vmPFC couples the secondary inducer event (the memory) to the somatic state reflecting “what it feels like” to be in a certain situation when one remembers it.
Once a somatic marker is formed and we are exposed to a similar stimulus, a change in the body’s periphery also triggers the initial recall of previously stored somatic markers. Signals from this activation are relayed to subcortical and cortical structures, notably the Insula and the somatosensory cortices. Once a somatic state is then produced, these same structures act on both a conscious and unconscious level to influence activity within brain regions involved in the generation of feelings and somatic states. These are the amygdala and ventromedial prefrontal cortex, as well as regions involved in working memory, ie. the dorsolateral prefrontal cortex and higher association cortices. Finally, somatic states influence activity within regions involved in motor and behavioral responses, either increasing or decreasing the likelihood an action will be taken.
Anterior Cingulate Cortex
emotional conflict resolution
A while ago, we wrote a blog post on the anterior cingulate cortex (see diagram below) where we looked at its role specifically in emotional conflict resolution.
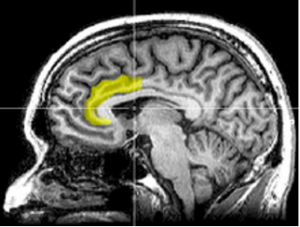
That discussion of emotional conflict was based on a 2003 paper by Rolls which offered a theoretical structure for emotion “in which motivational states are states in which instrumental goal-directed actions are performed to obtain rewards or avoid punishers, and emotional states are states that are elicited when the reward or punisher is or is not received.” This fits with our focus here on the cortical re-representation of limbic emotional processes integrated with symbolic logic or cognitive-based decisions and planning. It can be seen as part of an action-outcome reflection.
Of course, the anterior cingulate cortex is known to connect with the OFC and limbic areas in the amygdala, again making a complex circuit. In this action-outcome thinking, the posterior and mid-cingulate (not shown in the diagram above) have connections with the premotor area. They also get spatial and action inputs from the parietal cortex. The anterior cingulate may also be involved in affect-emotion regulation, particularly when it is challenging to do so. This will be an important topic later in this blog and throughout the blog series when we look at how a college student’s long-planned academic career path (e.g going to medical school) can be overturned by experience working in a neuroscience research laboratory, or not).
Dorsomedial Prefrontal Cortex
judging the mental states of others
As we wrote about in another blog, the dorsal medial prefrontal cortex, seen in the diagram below, seems to specialize in reading the mental states of others. In psychology, this is called the theory of mind – the idea that the person with whom you are interacting has a mind with plans and ideas that are indicated by their actions and emotional displays.
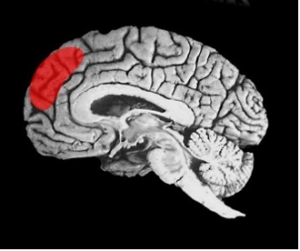
It just seems wise to try to understand another’s goals from their behaviors. This may be behind the idea of mirror neurons where what appear to be motor command neurons for complex actions will fire when the primate sees another perform that task. They seem to be particularly sensitive to the goal of a behavior, such as another person feeding themself (picking up, hand-to-mouth, etc.), rather than the just the simple movement. Some have suggested that mirror neurons could be involved, with other processes, in helping us to develop a high level process of empathy.
This dorsal medial prediction about the intentions of others extends to other predictions made in the world, such as whether or not a reward is coming depending on contextual circumstances. What is interesting, and discussed in that aforementioned blog, is that manipulating input from the dorsomedial prefrontal cortex to the nucleus accumbens can alter the firing of the accumbens neurons. It is then as though the perceived reward values themselves are altered at the limbic level by what the dorsomedial cortex is doing (planning).
Such trade-offs happen to us all the time. Consider a famous one where a reward for animals, if delayed in time, has to be made bigger to claim the same behavioral attention. The phenomenon is known as “reward or temporal discounting,” but its operation is well-known by college students (and others) as partying tonight instead of properly studying for the exam tomorrow. They would not do that if the exam were given at 11:00PM instead of the next morning. But with delay, it loses some control over immediate behavior.
Broad principle – coordination between cortical areas re-represent emotional processes
The above discussion of structures all have one thing in common: they are areas where neural processing in the limbic system connects to processing in the neocortex. As such, we regard them as outposts of the symbolic logic process that happens in the cortex and underlies our cognitive thinking. We think of them as part of the value brain for cognitive planning. As we have repeated, this process re-represents emotional functions in the limbic system in many forms so that they can cognitively become part of a student’s conscious plans for themselves. Complementing the facts and theories presented by a college education with an internship in their chosen field gives a student a feeling about a career path. Of course, there are many other examples of such cognitive-emotional integration that are important in other walks of life.
A testimony to that is the 2002 award of the Nobel Prize in economics to psychologist Daniel Kaneman who wrote the famous book, Thinking Fast and Slow. In this book, an always-on fast-judgemental emotional system of evaluation (system 1) accompanies a slower more deliberative cognitive system of evaluation (system 2) when typical decisions are made. The results of system 1 are often referred to as heuristics or “rule of thumb” insights into a decision that can bias the previously thought of cognitive basis. In a classic example, Kahneman refers to the way an experience ends can have an outside influence on evaluation. For example putting one’s finger for 15 seconds in painfully cold water results in an evaluation that is actually worse than if one does that but follows it with an additional 5 seconds of painful but not as painful cold water. Although all of the previous pain was in the second experience, the slightly better end causes subjects to rate the entire experience as less negative. The question for us all is where do these emotional or gut-level processes hurt or help in evaluations and subsequent decisions.
Conclusion
The more we understand about brain areas associated with implicit processing, based on limbic system input but re-represented in the cognitive symbolic neocortex the better. There are several reasons for this. First is that it raises awareness in the student and in the higher education institution of the importance of and need for such input from internships and other experiential activities outside the classroom transmission of facts and theories. Second, it reminds us of the need for careful consideration of and reflection on these experiences and other experiences a student has in executing their major and minors in pursuing a potential career path. Reflection is needed to better and more completely cognitively surface these reactions as well as to consider if and how to modify the student’s college plan. Finally, modern brain scanners are teaching us much more about how our brains work at all stages of life and one lesson learned is that in college time learning from experience is brain-natural and it makes a great complement to the academic study of that field in classes.